| Table of Contents | |
|
Review Article
| ||||||
| A combined approach of teaching head development using embryology and comparative anatomy | ||||||
| Melinda Danowitz1, Hong Zheng2, Adriana Guigova3, Nikos Solounias4 | ||||||
|
1BA, Medical Student, Anatomy, New York Institute of Technology College of Osteopathic Medicine, Old Westbury, NY, USA.
2DO, Resident, Vascular Surgery, Philadelphia College of Osteopathic Medicine, Philadelphia, PA, USA. 3DO, Fellow, Hematology/oncology, North Shore- Long Island Jewish, New Hyde Park, NY, USA. 4PhD, Professor, Anatomy, New York Institute of Technology College of Osteopathic Medicine, Old Westbury, NY, USA. | ||||||
| ||||||
|
[HTML Abstract]
[PDF Full Text]
[Print This Article]
[Similar article in Pumed] [Similar article in Google Scholar] |
| How to cite this article |
| Danowitz M, Zheng H, Guigova A, Solounias N. A combined approach of teaching head development using embryology and comparative anatomy. Edorium J Anat Embryo 2016;3:17–27. |
|
Abstract
|
|
Many aspects of human head embryology reflect its evolutionary development. The pharyngeal arches, a major component of head development, originally functioned in filter feeding and vascular exchange, which is why each arch has associated vasculature and muscles. The primitive tongue had few-associated muscles and was responsible for simple movements; the human tongue evolved post-otic somites that migrate to the tongue and develop the majority of the tongue musculature. These somites originate outside the tongue, and the motor innervation therefore differs from the general and special sensory innervation. In the primitive condition, the trapezius and sternocleidomastoid belonged to a single muscle group that were involved in gill movements; they separate into two muscles with the reduction of certain skeletal elements, but retain the same innervation. Examining the evolutionary changes of many structures allows for a greater understanding of the human embryology, and removes the need for memorization of seemingly complex processes. A link to comparative evolutionary anatomy provides context to the purpose and morphology of primitive structures, and clarifies several issues in human head development.
| |
| Keywords: Anatomy education, Embryology, Head and neck, Pharyngeal arches | |
|
Introduction
|
|
The development of the human head is of critical importance to the medical students, as a complete understanding is necessary to comprehend pathologies such as cleft lip and palate, Treacher Collins syndrome, and DiGeorge syndrome, among many others. These pathologies have been studied in mice in addition to humans, allowing for the establishment of genetic homologies [1] [2] [3] . The embryology of the human head is strongly related to the evolutionary history of vertebrate head and neck. We believe a link to comparative evolutionary anatomy is needed, as it provides context to the original purpose and development of early structures. An AAMC 2000 Graduation Survey revealed that 73% of respondents never took a premedical comparative anatomy course [4]. The American Association of Medical Colleges and the Howard Hughes Medical Institute recommended that evolutionary biology should be included in medical education [5]. Without an understanding of the evolutionary history, head embryology can be an overly complex topic that involves a great deal of memorization. Furthermore, anomalous anatomical presentations often reflect the typical morphology in more primitive animals; studies of human biliary tract and limb musculature variations revealed that the atypical human structures resembled the normal condition seen in other animals [6][7]. The study of evolutionary biology relates not only to anatomy and embryology, but also to medicine in general. At the molecular level, evolution helps biologists study genetic mutations that allow viruses to infect multiple species, or how pathogens gain virulence [8]. Darwinian medicine also facilitates comprehension about natural selection, and which aspects of the human body are vulnerable to, or protected against disease [5]. Natural selection explains malarial resistance in sickle-cell trait carriers, and can potentially clarify associations between breast cancer and birth control [9]. Studies of medical education curricula in the UK as well as in North America demonstrated an overall need for the incorporation of evolutionary biology into medical courses [10][11]. The evolutionary origin of the skull, face, and neck has been extensively studied from a scientific standpoint [12] [13][14][15]. For example, studies involving transplanting avian embryonic cells and tracking the differentiation of cranial tissues gave insight into the fate of mesodermal and neural crest cells [16]. We aim to link human head embryology with evolutionary history at a level understandable for medical students. We, therefore, provide a brief description of the evolutionary history of each process involved in head development, to provide context to the embryonic structures and ultimate derivatives, and to help students understand why the human anatomy develops from the early embryo the way it does. We currently provide a comprehensive summary of the development of the human head. The aim for this article is to be used as a supplement to the various medical embryology textbooks. We believe the abbreviated yet comprehensive summary of head and neck embryology, supplemented with a section describing the comparative evolutionary anatomy, will enable medical students to better understand the development of the head. |
|
The General Development of the Human Head
|
|
The head is formed by the interaction of the three basic tissues: ectoderm, mesoderm and endoderm. When the head is fully differentiated, its parts are the result of combined tissues from all three germ layers. The sections of this study are the components of the head. Each is treated here separately but they all interact and form an integrated whole. Many of the interactions take place simultaneously. |
|
Brain, Heart, Pharynx, and Notochord
|
|
The head begins to form with interactions between four basic early structures: brain, heart, pharynx and notochord (Figure 1). The brain forms as an involuting set of neural folds from the rostral end of the neural tube. The original embryological external surface of the neural tube will ultimately form the internal folds and ventricles of the brain. The early heart develops anterior to the brain. As development progresses, the heart slowly swings posteriorly and many of its arteries will become incorporated into the pharynx as the pharyngeal arch arteries (Figure 1). Thus, the proximal heart circulation is closely related to the head. Pharyngeal, or branchial arches are 6 units of complex tissues aligned in a sequence. The pharynx has pouches and clefts along its length. A cleft-pouch unit alternates with an arch. The clefts (externally) and the pouches (internally) correspond to each other and are separated by a membrane (Figure 2). Cleft 1 is the only cleft that persists in humans; the first cleft and pouch fuse to form the tympanic membrane of the ear. All other clefts are obliterated by the down growth of the platysma muscle from arch 2. Each pharyngeal arch includes an associated artery, vein, a branchiomeric cranial nerve, muscles, cartilage, and neural crest cells. Pharyngeal arch 1 is the only arch that contains two processes (maxillary and mandibular processes) whose derivatives are ultimately connected by a hinge joint (temporomandibular joint) (Figure 1). The upper jaw (maxilla) is not derived from the first arch pharyngeal cartilages. The pharynx is an internal space around which the arches appear to wrap around. The arches and the cleft-pouch units constitute the wall of the pharynx. The median area is the pharyngeal floor, which constitutes the mucosal surface, but not the internal muscles, of the tongue [17]. The anterior part of the pharynx connects to an external tunnel, the stomodeum, which forms under the developing brain. The stomodeum constructs the oral and nasal cavity (initially undivided regions), and is lined by an ectodermal membrane (Figure 2). The stomodeum is separated from the pharynx distally by the double-lined oropharyngeal membrane, which is both ectodermal and endodermal. The contact between the stomodeum and pharynx is one of two areas in the body where ectoderm directly touches endoderm; this is positioned near the palatine tonsils. The oropharyngeal membrane will open to form a patent pathway between the nasal and oral cavities, and the pharynx. Inside the primitive pharynx there is endodermal lining (mucosa) which does not extend to the stomodeum. The anterior lining of the oral cavity is ectodermal mucosa. The two mucosae eventually become indistinguishable. At the roof of the stomodeum is Rathke's pouch, which penetrates the head and will contribute to the formation of the anterior pituitary [18]. |
|
|
|
|
|
|
|
Cranial Nerves
|
|
Historically the 12 cranial nerves were counted in the sequence as they exit the brain; I is the first and XII is the last (Figure 3). The numbers have been retained in literature but their functions were discovered more recently. The numeric sequence of the cranial nerves does not reflect their function. Three types of cranial nerves are involved in the function of the head: (1) Cranial nerves associated with placodes. These are special sensory nerves that innervate the nose, the eye, and the ear, respectively (CN I, II, and VIII). (2) Cranial nerves associated with the head somites and somitomeres, which are strictly motor in function (CN III, IV, VI, XI, and XII). There are three preotic somites, each innervated by its own motor nerve (CN III, IV, VI), creating the extra-ocular muscles. There are two groups of postotic somites, creating the sternocleidomastoid and trapezius (CN XI), and the intrinsic muscles of the tongue (CN XII). (3) Cranial nerves of the pharyngeal arches (or branchial arches): the branchiomeric nerves (CN V, VII, IX and X). This group of cranial nerves has mixed modalities; they innervate the pharyngeal arch musculature and the larynx and have motor, sensory, and glandular secretory functions [19]. Two additional cranial nerves have been established and studied in fishes and in some other vertebrates. The nervus terminalis is associated with the olfactory nerve, and has been demonstrated in fishes to relate closely to luteinizing-hormone releasing hormone, but the presence and function in humans is not well known [20][21][22]. The nervus intermedius is better established in humans, and relates to sensory and parasympathetics of the facial nerve [23][24]. This nerve is potentially implicated in pathologies such as nervus intermedius or geniculate neuralgia and cluster headaches [25][26][27]. |
|
|
|
Head Forms Outside The Coelom
|
|
In general, the body can be divided into two types of regions: the musculoskeletal system, and the core viscera containing the organs. The organs are surrounded by spaces and membranes (pleural and peritoneal), and are enclosed by the musculoskeletal system, which forms a surrounding tube. The organs do not contact the surrounding muscles and bones, but are instead contained within spaces that are lined by membranes termed the pleural and peritoneal membranes of the coeloms. The organization of the head and neck differs from that of the remainder of the body, where the essential organs (eyes, tongue, larynx, etc.) are not enclosed by coeloms. The head and neck viscera therefore contain no visceral nor parietal peritonea [28]. Evolutionary link The specialized organs of the head evolved prior to the evolution of the coelom. The sensory organs now concentrated in the head were surface structures that allowed the organism to directly interact with its surroundings. In fishes, these specialized structures, such as olfactory and vision organs appeared in the anterior part of the body, as this is the region that first contacted the changing environment, and therefore the organism could respond rapidly to its surroundings. Enclosure of these sensory organs within an internal coelom would prevent their direct contact with the environment and therefore impede their function [30]. |
|
Neural Crest Cells
|
|
Neural crest cells are an essential component to the development of the head. Neural crest forms at the edge of the infolding neuroectodermal brain tube and later detaches and migrates ventrolaterally (Figure 4). This is a separate tissue, which plays a huge inductive role in the formation of the head and of many other parts of the body. The neural crest descends in complex fashion from its original location and becomes involved in differentiation of structures throughout the body, especially of the head mesoderm (or mesenchyme) and the head ectoderm [19]. It is not possible for the mesoderm to form structures in the head without the organizing forces of the neural crest. For this reason, many textbooks refer to neural crest derivatives as ectodermal, rather than mesodermal in origin. The neural crest directs the mesodermal differentiation, and creates the fascial lattices that enclose all these mesodermal structures. Neural crest builds the scaffolding for the mesenchyme to form head structures. The neural crest flows downward from its original connection with the neural tube to interact with the mesoderm and the ectoderm of the head (Figure 4) [17]. The neural crest directs the connection of the head with the axial skeleton (e.g., neck and thorax). Evolutionary link |
|
|
|
The Developing Brain
|
|
The brain begins as a tube formed by the infolding of the neural surface. Regions begin to differentiate into segments that relate to discrete functions (Figure 4). The rostral brain begins as the prosencephalon, which divides into the telencephalon and diencephalon. The telencephalon creates the cerebral hemispheres. The diencephalon consists of four prosomeres, forming the thalamus, epithalamus, hypothalamus, pineal gland, and posterior pituitary gland. The mesencephalon forms the midbrain. Caudal to this is the rhombencephalon, or hindbrain, which divides into the metencephalon and myelencephalon. The rhombencephalon forms eight constrictions, which are termed rhombomeres, forming the various parts of the adult hindbrain [32]. Evolutionary link |
|
|
|
Skull Bones and Related Structures
|
|
Two types of ossifications occur in the skull; intramembranous and endochondral. During intramembranous ossification, bones develop directly from mesoderm with no cartilaginous precursor. During endochondral ossification, a cartilaginous precursor is subsequently replaced by osteocytes. Both types of bone are mesodermal, with a strong inductive influence form the neural crest, which forms the fascial lattices of the head [17]. The four major components of the skull include the dermal skull bones (intramembranous), splanchnocranium (intramembranous), neurocranium (endochondral), and the branchial arch cartilages (endochondral) (Figure 6). (1) The intramembranous bones are mesodermal and differentiated by the neural crest cells. They ossify and from a dome that fuses to the underlying neurocranium. These include the nasal, frontal, zygomatic, lacrimal, palatine, vomer, and parietal bones. (2) The splanchnocranium, or viscerocranium, are intramembranous bones that cover pharyngeal cartilages. These include the maxilla and mandible. (3) The neurocranium is a bowl shaped structure, formed initially by mesenchymal endochondral cartilages that later ossify. The endochondral skull bones include the turbinates, ethmoid, and basioccipital bones. These two fuse and form one eventually ossified spherical skull. The neurocranium also includes five parachordal somites, which are carried into the head with the notochord; these form the bony area around the foramen magnum. The sphenoid, the temporal and the occipital bones form by a combination of the intramembranous and endochondral components. (4) The branchial cartilages are components of the pharyngeal arches. The alisphenoid and incus ossify from the maxillary cartilage, and the malleus ossifies from Meckel's cartilage; both sets of cartilages derive from pharyngeal arch 1. The stapes, styloid process, and the lesser horn and portion of the body of the hyoid ossify from Reichert's cartilage of the second arch. The remainder of the hyoid derives from third arch cartilage, and the thyroid and laryngeal cartilages derive from the 4th and 6th arches [34]. Evolutionary link |
|
|
|
|
|
|
|
Head Muscles
|
|
Around the skull there are muscles, which are of two types: those from the branchial apparatus and those from the head somites (Figure 8). Ultimately, all the muscles are mesenchymal but their differentiation is influenced by the neural crest. Many of the head muscles are derived from the branchial arches, including the muscles of mastication (arch 1), muscles of facial expression (arch 2), stylopharyngeus (arch 3), constrictors of the pharynx (arch 4), and intrinsic muscles of the larynx (arch 6). Muscles from head somites are from two regions, preotic and postotic, with the ear region being used as a landmark for differentiation. There are three preotic somites, and each is associated with a motor cranial nerve. The first preotic somite sub-divides and creates the four extra-ocular muscles innervated by the oculomotor nerve (inferior oblique, medial rectus, superior rectus, inferior rectus). The second develops the muscle innervated by the trochlear nerve (superior oblique). The third preotic somite creates the extra-ocular muscle innervated by the abducens nerve (lateral rectus). There are two muscle groups derived from the postotic somites. The intrinsic muscles of the tongue, innervated by the motor hypoglossal nerve, are derived from the first four postotic somites. The two muscles innervated by the spinal accessory nerve, the sternocleidomastoid and trapezius, are derived from postotic somites 5–7. There is a cranial boundary that is defined by Hox genes, and somites caudal to this boundary are associated with spinal nerves and contribute to the formation of hypaxial and epaxial musculature of the body [18]. Evolutionary link |
|
|
|
Head Placodes
|
|
As the head is developing, distinct ectodermal discs called placodes invaginate and sink into the head bilaterally. There are two different types of placodes: neurogenic and non-neurogenic (Figure 3). (1) The neurogenic placodes will become the ganglia of cranial nerves, and also include the otic and olfactory placodes. The otic, or auditory placode forms the cochlea and vestibular organ of the ear, and the olfactory placode gives rise to the olfactory epithelium in the nose. (2) The non-neurogenic placode develops aspects of the eye. The formation of the eye results from the interaction between the lens placode and the optic cup, which is an extension of the developing brain [17]. Evolutionary link |
|
Nasal and Oral Regions
|
|
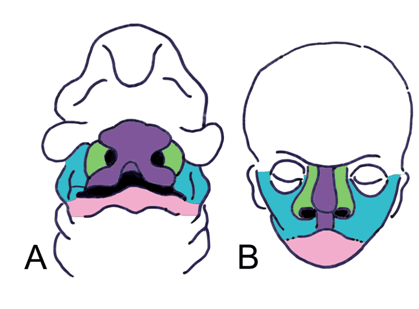
The primitive face commences as a frontonasal prominence (swelling), and a pair of maxillary prominences, and mandibular prominences, all influenced by neural crest cells (Figure 9). These swellings surround the stomodeum and the invaginated nasal placode. The swellings are mesodermal and ectodermal and form a complex surface external to the developing skull bones. The maxillary prominences grow anteromedially, and concurrently, ectodermal olfactory placodes enlarge and divide the frontonasal prominences into lateral and medial nasal processes. The nasolacrimal groove separates the lateral nasal processes from the maxillary prominences. The groove closes to form the nasolacrimal duct. The medial nasal processes fuse at midline to develop the bridge and septum of the nose, and expands laterally to form the intermaxillary process. The fusion of the maxillary processes with the intermaxillary process at the median plane separates the nasal and oral cavities and creates the primary palate, located anterior to the incisive canal. The external fusion forms the philtrum. The secondary palate (hard palate) is formed from medial extensions of the maxillary prominences, termed the palatine shelves. The fusion and interaction of these facial prominences and swellings create the external surface and cartilaginous features of the face [35]. The muscles of arch 2 migrate external to these prominences to form the face muscles. Evolutionary link |
|
|
|
Development of The Tongue
|
|
The tongue commences as a median swelling formed from the first pharyngeal arch, which is subsequently overgrown by two lateral swellings, also from the first arch (Figure 10). This forms the mucosa of the anterior two thirds of the tongue. The second arch forms the copula, which is a midline swelling that becomes overgrown largely by the hypopharyngeal eminence of the third arch, which creates the surface of the posterior one third of the tongue. A small contribution of the fourth arch creates the posterior-most surface of the tongue. The terminal sulcus delineates the contributions of the first and third arches. The ventral surface of the tongue initially is connected to the floor of the mouth. The anterior aspect of this connection regresses, and the posterior portion persists, forming the frenulum. Myotomes from occipital somites migrate inside the tongue, to form the musculature. The innervation of the tongue is consistent with the embryologic origin; the musculature is innervated by the hypoglossal nerve (occipital somites), sensory to the anterior 2/3 of the tongue is innervated by branches of the trigeminal nerve (first arch), sensory to the posterior 1/3 of the tongue is derived from the glossopharyngeal nerve (third arch), sensory to the posterior-most region of the tongue is innervated by the vagus nerve (fourth arch). Special sensory (taste) is derived from chorda tympani of the facial nerve (anterior 2/3), and from the glossopharyngeal nerve (posterior 1/3) [19]. Evolutionary link |
|
|
|
Glands
|
|
The thyroid originates as an endodermal mass at the apex of the foramen cecum of the tongue. This descends down the neck attached to the thyroglossal duct, and ultimately attaches inferior to the cricoid cartilage. Parafollicular C cells of the thyroid are derived from migrating neural crest cells. The salivary glands develop based on complex interactions between epithelial and mesenchymal cells. The palatine tonsil derives from the endodermal pouch of the second pharyngeal arch. The thymus develops from endoderm of the third pharyngeal pouch, and becomes infiltrated by neural crest cells, creating the septa and capsule of the immune gland. The inferior parathyroids are formed by the pouch of the third pharyngeal arch, and they migrate caudal to the superior parathyroids, which derive from the pouch of the fourth pharyngeal pouch [19]. Evolutionary link |
|
Conclusion
|
|
Studying comparative anatomy and evolution helps put human embryology into perspective, and often explains the resulting anatomical structures. Many morphological features reflect the evolutionary development, and familiarity of these processes reduces the need for memorization, and allows for a greater understanding of the embryology and anatomy. We believe reviews of each system of embryology supplemented with the comparative evolution will help students better understand human development and facilitate long-term retention of knowledge. |
|
Acknowledgements
|
|
We thank the Anatomy Department at New York Institute of Technology College of Osteopathic Medicine. |
|
References
|
|
|
[HTML Abstract]
[PDF Full Text]
|
|
Author Contributions
Melinda Danowitz – Substantial contributions to conception and design, Acquisition of data, Analysis and interpretation of data, Drafting the article, Revising it critically for important intellectual content, Final approval of the version to be published Hong Zheng – Analysis and interpretation of data, Revising it critically for important intellectual content, Final approval of the version to be published Adriana Guigova – Analysis and interpretation of data, Revising it critically for important intellectual content, Final approval of the version to be published Nikos Solounias – Substantial contributions to conception and design, Acquisition of data, Analysis and interpretation of data, Drafting the article, Revising it critically for important intellectual content, Final approval of the version to be published |
|
Guarantor of submission
The corresponding author is the guarantor of submission. |
|
Source of support
None |
|
Conflict of interest
Authors declare no conflict of interest. |
|
Copyright
© 2016 Melinda Danowitz et al. This article is distributed under the terms of Creative Commons Attribution License which permits unrestricted use, distribution and reproduction in any medium provided the original author(s) and original publisher are properly credited. Please see the copyright policy on the journal website for more information. |
|
|